|
А.В.Никитин Считывание и обработка информации ДНК.
К этой теме я обращался уже не раз [9, 10] и с разных
сторон. Но как потом оказывалось, изложение затрагивало только часть процесса,
и далеко не полную. Что-то важное все время упускалось. Очень уж сложным
выглядит описание этого процесса в материалах специалистов. Специфика языка и
биологическая терминология не позволяет быстро понять и осознать смысл
прочитанного. У специалистов свой язык описания процессов. Нам его без
перевода понять трудно. Но, начинать надо все же, с оригинала… Да простят меня читатели за большие цитаты. Но иначе
было невозможно показать, как выглядят процессы считывания и использования
информации ДНК на языке профессионалов. Вот и пришлось давать такие длинные
цитаты, а потом комментировать их уже более понятным для нас дилетантов
языком. Конечно, говорить о полноте изложения и подборки
материалов не приходится. Это только очередной подход с тех позиций понимания,
которые открываются по мере изучения этой темы. Зарисовка на тему, снова и с
самого начала… Эукариоты и прокариоты.
Биологи делят клетки на два больших надцарства [6]:
эукариоты и прокариоты. К первому надцарству клеток относятся и наши клетки. К
надцарству прокариотов отнесены
простейшие. Вирусы, бактерии… Вот примерно так: «Все организмы, имеющие клеточное строение, делятся на
две группы: предъядерные (прокариоты) и ядерные (эукариоты). Клетки прокариот, к которым относятся бактерии, в
отличие от эукариот, имеют относительно простое строение. В прокариотической
клетке нет организованного ядра, в ней содержится только одна хромосома,
которая не отделена от остальной части клетки мембраной, а лежит
непосредственно в цитоплазме. Однако в ней также записана вся наследственная
информация бактериальной клетки. Цитоплазма прокариот по сравнению с цитоплазмой
эукариотических клеток значительно беднее по составу структур. Там находятся
многочисленные более мелкие, чем в клетках эукариот, рибосомы. Функциональную
роль митохондрий и хлоропластов в клетках прокариот выполняют специальные,
довольно просто организованные мембранные складки.
По строению различные эукариотические клетки сходны. Но
наряду со сходством между клетками организмов различных царств живой природы
имеются заметные отличия. Они касаются как структурных, так и биохимических
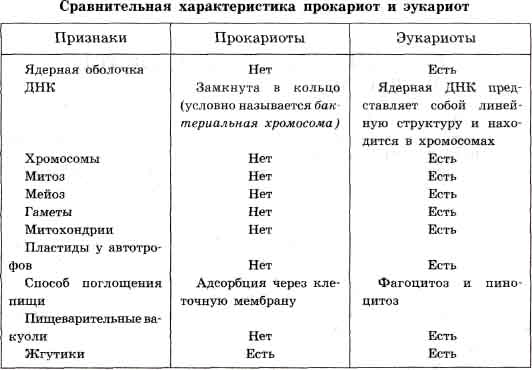
особенностей». [5] «…высшие животные, растения и грибы состоят из клеток,
в которых находится одно ядро. Оно имеет форму шара с диаметром от 3 до 10
мкм (рис. 1). [5].
Рис.1. Ядро клетки. «…в ядре клеток заключены хромосомы, которые содержат ДНК
- хранилище наследственной информации. Этим определяется ведущая роль
клеточного ядра в наследственности».[5] С основным различием клеток разобрались. Идем дальше. Основные данные.
Теперь посмотрим, что такое ДНК и РНК. ДНК, это то, что составляет хромосомы. Это постоянная
память клетки. То, что создает её наследственную основу. Хромосома –
хранилище информации. ДНК – отдельный информационный массив из этого
хранилища. Но, очень большой массив. Да и хорошо защищенный. Чтобы воспользоваться информацией с ДНК необходимо
сделать несколько действий: Спираль ДНК разворачивается и разрезается вдоль
оси. Потом каждая одиночная последовательность ДНК копируется. Так получается
РНК. По частям – генам. Получается мРНК-предшественник. И только потом из
предшественника получается форма мРНК, пригодная для трансляции, т.е. синтеза
белка. Стандартная мРНК, это последняя стадия в длинной цепи
доработки копии ДНК, подготовленная
для начала считывания информации. Этот термин биологи не применяют.
Но, мы его введем. Для эукариотов. К
стандартной мРНК мы еще вернемся… А пока основные данные о ДНК и РНК: «Как известно, оснований, которыми различаются
нуклеотиды, всего четыре. В РНК это аденин (A), гуанин (G), цитозин (C) и урацил (U) (T-тимин в ДНК), а обычных аминокислот, входящих в белки, - 20.
Следовательно, задача сводится к тому, чтобы четырьмя основаниями записать
двадцать аминокислот. И отсюда следует, что код должен быть не менее чем
триплетным, поскольку по одному основанию и даже по два (4 x 4 = 16) недостаточно, а по три даже много (4 x4 x 4 = 64). Сколько же кодонов из
64 имеют смысл, а какие бессмысленны? Соответствует ли каждой аминокислоте
один или несколько кодонов? Ответы на эти вопросы были получены к 1965 году,
когда генетический код был полностью расшифрован». [2] «…код триплетен - каждая кодирующая единица-кодон
состоит из трех нуклеотидов. В каждом гене триплеты
считываются с фиксированной точки, в одном направлении и без запятых, то есть
кодоны ничем не отделены друг от друга». [2] «Сначала информация, записанная в виде чередования
дезоксирибонуклеотидов на одной из двух комплементарных цепей в ДНК гена,
переписывается на одноцепочечную молекулу информационной рибонуклеиновой
кислоты - иРНК (она же мРНК от англ. messenger - переносчик). Это процесс транскрипции. На
следующем этапе по матрице иРНК строится последовательность аминокислотных
остатков полипептида. Тем самым создается первичная структура будущей
молекулы белка. Это процесс трансляции». [2] «Синтез белка или трансляция - это центральное событие в
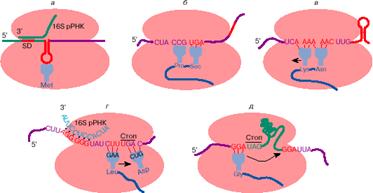
жизни клетки (рис.
2). Само
представление о дискретных единицах генетической информации существует
благодаря трансляции нуклеотидных последовательностей мРНК, ограниченных
кодонами: инициатором и терминатором. Считывание таких последовательностей
приводит к появлению в клетке дискретных молекул белка. Этот процесс
обставлен в клетке весьма сложно (см., например, [6]). В трансляции каждого
гена участвуют несколько сот разных молекул белков и нуклеиновых кислот». [2] Таблицы соответствия кодонов и аминокислот можно
посмотреть в [1,2]. Выяснили мы пока только то, что генетики и биологи
рассматривают ДНК с позиций
копирования (транскрипции) с получением копии – РНК и процесса
получения сборки белка (трансляции). Ну,
хоть с терминологией начали разбираться… Основания и правила…
Теперь посмотрим, что послужило основанием для формирования постулатов
считывания ДНК, принятых в генетике: «В 1961 году Ф. Крик и С. Бреннер экспериментально
показали, что делеция (вырезание) одного нуклеотида, дающая мутантный
фенотип, может быть скомпенсирована вставкой нуклеотида вблизи делеции. Этот
результат можно было объяснить предположив, что при делеции нуклеотида
происходит сдвиг рамки считывания за местом делеции, и это приводит к полному
изменению смысла всей последующей информации; при вставке одного нуклеотида
вблизи места делеции происходит восстановление первоначальной рамки
считывания и первоначального смысла закодированной информации. Таким образом,
описанные эксперименты доказали, что генетический код не содержит запятых. В
опытах с делециями и вставками Крик, Барнет, Бреннер и Уотс-Тобин (1961)
также подтвердили, что код является триплетным или кратен трем, поскольку три
делеции или три вставки нуклеотидов давали нормальный фенотип. То есть, если вырезать один нуклеотид … и тут же рядом
вставить, то … ничего не изменится. Это подтверждает отсутствие запятых…, а
так же и триплетность кода. А вот опыты Г. Виттманна - это серьезнее. Если учесть то, что сказано далее о
нечувствительности трансляции к третьему элементу кода триплета, то… этот постулат не совсем верен.
Неперекрываемость генетического кода не подтверждается. Коды могут перекрываться, но мы это далеко
не всегда видим. Таким образом, подтвержденными можно считать только
триплетность и непрерывность кода ДНК и мРНК. Вот теперь можно считывать информацию с ДНК и мРНК.
Здесь правила примерно одинаковы. При
этом: «…первый же встреченный на иРНК кодон AUG (Met) задает фазу последующего
считывания троек, то есть служит той самой фиксированной точкой, с которой
начинается считывание. Любой последующий AUG просто кодирует Met. В конце гена обязательно
стоит UAA, или UAG, или UGA, а то и два нонсенса подряд». [2] Видимо, это обычный порядок считывания. Начало и конец
считывания установлен. Оказывается, мы давно знаем это. Стандартная мРНК.
Как мы уже говорили,
процесс получения стандартной мРНК достаточно длинный. Вот как это
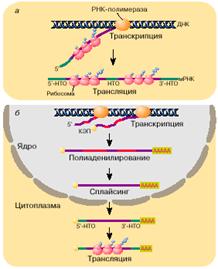
происходит: «Эукариотические мРНК … Их транскрипция и
трансляция пространственно разобщены. Транскрипция протекает в ядре, а
трансляция - в цитоплазме (рис. 2, б ).
Эукариотические мРНК синтезируются в виде предшественников и проходят в своем
биогенезе стадию довольно сложного созревания, или процессинга. Процессинг
включает в себя: 1) кэпирование 5'-конца, заключающееся в присоединении к
этому концу мРНК так называемой шапочки (кэп-структуры), 2)
полиаденилирование 3'-конца и, наконец, 3) сплайсинг - вырезание протяженных
внутренних участков мРНК, так называемых интронов, и ковалентное
воссоединение оставшихся фрагментов (экзонов) через обычную фосфодиэфирную
связь…». [1].
Процессинг имеет
целью сформировать различные области мРНК в нужном для трансляции порядке. «Те части молекулы мРНК, в которых закодированы белки,
называются транслируемыми областями. Однако помимо транслируемых областей в
мРНК имеются достаточно протяженные последовательности, не кодирующие белок.
Общая длина этих нетранслируемых областей порой может достигать или даже
превышать длину транслируемых областей. [1] Перестановки и удаления фрагментов мРНК выполняются на этапе сплайсинга: «Сплайсинг (от англ. splice — сращивать или склеивать концы чего-либо) — процесс вырезания определенных нуклеотидных
последовательностей из молекул РНК и соединения
последовательностей, сохраняющихся в «зрелой» молекуле, в ходе процессинга
РНК. Наиболее часто этот процесс встречается при созревании информационной
РНК (мРНК) у эукариот, при этом путём биохимических реакций с участием РНК и белков из мРНК
удаляются участки, не кодирующие белок (интроны) и соединяются друг с другом кодирующие аминокислотную
последовательность участки — экзоны. Таким образом незрелая пре-мРНК превращается в зрелую мРНК, с которой
считываются (транслируются) белки клетки. Большинство генов прокариот,
кодирующих белки, не имеют интронов, поэтому у них сплайсинг пре-мРНК
встречается редко. У представителей эукариот, бактерий и архей встречается
также сплайсинг транспортных РНК (тРНК)[1] и других некодирующих РНК». [8] Нетранслируемые области внутри мРНК, как мы поняли,
называются интронами. Попробуем выяснить, что из себя представляют интроны,
которые удалятся при сплайсинге стандартной мРНК: «Существуют две альтернативные теории, объясняющие
происхождение и эволюцию сплайсосомных интронов: так называемые теории ранних
интронов (РИ) и поздних интронов (ПИ). Теория РИ утверждает, что
многочисленные интроны присутствовали в общих предках эу- и прокариот и,
соответственно, интроны являются очень старыми структурами. Согласно этой
модели, интроны были потеряны из генома прокариот. Также она предполагает,
что ранние интроны способствовали рекомбинации экзонов, представляющих домены белков. ПИ утверждает, что интроны появились в генах относительно недавно и
были инсертированы (вставлены) в геном после разделения организмов на про- и эукариоты. Эта модель основывается
на наблюдении, что сплайсосомные интроны есть только у эукариот». [7] «Почти все эукариотические ядерные интроны начинаются с GU и оканчиваются AG (правило AG-GU)». [7]
Рис.3.Схема нуклеотидной
последовательности пре-мРНК гена CDK4 человека. Большую часть последовательности занимают интроны (показаны
серым цветом) Для прокариотов
значимость интронов установлена более точно: «В
прокариотических полицистронных мРНК имеются также внутренние межцистронные
нетранслируемые области, располагающиеся между транслируемыми областями.
Наряду с информацией о последовательности аминокислот в белке молекулы мРНК
содержат информацию, определяющую их поведение в клетке (активность и время жизни,
внутриклеточное распределение). Эта информация находится в основном в
нетранслируемых областях мРНК». [1]. Вот так. Что такое интроны – узнали мало, но
выяснилось много нового… Если рассматривать мРНК-предшественник, как общую
информацию о том, или ином белке, то экзон
надо рассматривать, как исполнительную программу производства белка на рибосоме, а интрон – дополнительная информация об
этом. Например, что с чем сшивать и в каком порядке, как настроить рибосому
на этот процесс, перечень дополнительных команд, и т.д. Интроны, как раз и есть кодированная запись команд и
синхронизации всего производства белка
- главное в этом процессе. Очень возможно, что интроны обрабатываются
и частично вставляются в области 5' и 3', как дополнительная информация для
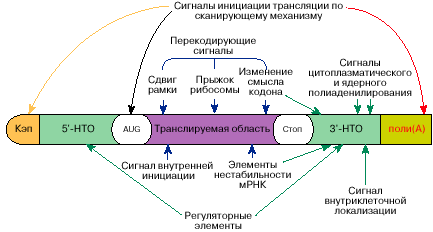
считывающего устройства рибосомы. В сформированной для трансляции мРНК: Нетранслируемые области находятся на обоих концах
молекул мРНК и соответственно называются 5'- и 3'-НТО». [1] Полиаденилирование 3'-конца закодировано в самой мРНК. Потому, что: «Около половины мРНК эукариот полиаденилируются на
3'-конце во время процессинга в клеточном ядре. Сигналом ядерного
полиаденилирования 3'-конца служит последовательность AAUAAA, расположенная за 10-20 нуклеотидов от этого конца».
[1]. «Такой последовательностью у амфибий является (U)6AUAAAG. Поли(А)-хвост на мРНК
узнается особым поли(А)-связывающим белком, который участвует в инициации
трансляции мРНК по кэп-зависимому механизму». [1]. И получилась мРНК, пригодная для сборки белка. Наша - такая же, как на
рис 2.(б). Теперь посмотрим на рис.4. Это и есть стандартная форма
мРНК. Ничего не напоминает? А ведь весьма похоже на строку из
памяти компьютера. Составляющие почти те же… Только чуть сложнее. Разрешающий код, потом код начала
строки и программа действий, «старт-код», собственно информационная часть,
«стоп-код», проверочные и контрольные суммы, дополнительная информация о
порядке выполнения действий, и, наконец, код окончания строки.
«Зрелые эукариотические мРНК, как правило,
моноцистронны и кодируют только одну полипептидную цепь». [1] Таким образом, можно сказать, что стандартизация мРНК
развивалась вместе с развитием клетки.
Вариации на тему.
Теперь заметим, как объясняют специалисты
множественность триплетных кодов для одной и той же аминокислоты: «…для кодирования большинства аминокислот существенны
два первых основания, а третье может быть любым. Следовательно, мутации -
замены оснований в третьем положении многих кодонов просто не будут
проявляться. Кроме того, ограниченные возможности проявления имеют и мутации,
приводящие к замене полярного остатка на полярный или неполярного на
неполярный, поскольку они часто близки по своим физико-химическим свойствам.
Если такие мутации и проявляются, то проявляются нечетко, то есть мутантный
белок не полностью утрачивает свою активность, а лишь частично. Так могли
возникать в эволюции так называемые полипептиды-синонимы, имеющие одинаковую
укладку и ферментативную активность, но разную первичную структуру.
Получается, что генетический код обладает высоким уровнем помехоустойчивости
в том, что касается миссенс-мутаций, или мутаций, изменяющих смысл кодонов.
Чаще всего проявляются те миссенс-мутации, которые приводят к заменам
полярных остатков на неполярные и наоборот». [2] А дальше и совсем замечательно: «Инициирующий кодон узнается только в определенном контексте.
Если мы зададим вопрос, можно ли, имея перед собой последовательность
нуклеотидов какой-либо мРНК, таблицу генетического кода и зная, что
трансляция мРНК идет в направлении от 5'- к 3'-концу, а белковая цепочка
растет от N-конца к C-концу, написать последовательность аминокислот белка,
закодированного в этой мРНК, то будем вынуждены ответить на поставленный
вопрос отрицательно». [1]. То есть, записано одно, а транслируется – другое. Для
правильного считывания надо получить дополнительную информацию. И куда-то её
вовремя внести. Только с учетом этой информации процесс чтения пойдет в
нужном направлении: «У эукариот инициация происходит, как уже говорилось,
чаще всего с первого AUG, однако только в том случае,
если этот AUG находится в оптимальном контексте:
за два нуклеотида до него обязательно должен находиться пурин (A или G), а непосредственно за ним
должен следовать G. На эффективность инициации у эукариот определенное
влияние могут оказывать также нуклеотиды и в других положениях вблизи инициирующего
кодона. Самым оптимальным для узнавания инициирующего кодона у млекопитающих
считается следующее его окружение: GCCGCCA / GCCAUGGA / CU ». [1]. «Если первый AUG в
эукариотической мРНК находится не в оптимальном контексте, он пропускается и
инициация начинается со следующего AUG. Для такой инициации очень важно также наличие
кэп-структуры на 5'-конце мРНК и, как ни странно, поли(А) последовательности
на противоположном конце молекулы. Кэп-структура и поли(А) последовательность
узнаются специфическими белками, которые также необходимы для инициации. При таком способе инициации трансляции у эукариот
последовательность мРНК как бы просматривается (сканируется) с начала мРНК
(от ее кэп-структуры) для поиска кодона AUG в оптимальном контексте. Такая инициация получила название
"кэп-зависимая инициация по сканирующему механизму". Следует, однако, заметить, что на некоторых мРНК
эукариот инициация происходит не путем сканирования мРНК с 5'-конца, а за
счет непосредственного узнавания определенного внутреннего AUG. Для такого узнавания требуется весьма протяженная
предшествующая последовательность мРНК. Эта последовательность узнается особыми клеточными
белками, которые способствуют инициации трансляции по механизму
"внутренней инициации". По такому механизму инициируется трансляция
на многих вирусных РНК, а также на некоторых клеточных мРНК, кодирующих очень
важные регуляторные белки, например факторы роста фибробластов. Содержание
этих белков обычно очень мало, а увеличение их количества в клетке может
сопровождаться трансформацией клеток в раковые. Некоторые вирусы, генетическая информация которых
считывается по механизму внутренней инициации трансляции, способны выключить
инициацию трансляции клеточных мРНК по сканирующему механизму и, таким
образом, переключать белоксинтезирующий аппарат клетки на синтез собственных
белков». [1]. Нормально, как в нашем родном русском языке.
Отступлений от правил больше, чем самих правил. Оказывается, что для
правильной трансляции белка надо выполнить множество дополнительных условий. Но, специалисты
по вычислительной технике сразу поняли - в чем дело. Если рассматривать
процесс трасляции с этой технической стороны, то под контекстом понимаются
нужные разрешающие команды на нужные управляющие входы считывающего
устройства и переход к чтению того фрагмента из 5' или 3', который отвечает
за данный участок трансляции. Потом в соответствии с этим скорректировать
свои действия. «Избирательное влияние на активность мРНК в трансляции
оказывают специфические регуляторные белки или специальные регуляторные РНК.
Эти белки или РНК проявляют свое действие, связываясь со специфическими
последовательностями или структурами в мРНК, которые называются регуляторными
элементами. В большинстве случаев регуляторные элементы располагаются в
5'-НТО вблизи инициирующего кодона. Однако в некоторых случаях регулярные
элементы могут быть на значительном расстоянии от инициирующего кодона, в том
числе в 3'-НТО. Связываясь с мРНК вблизи инициирующего кодона,
регуляторные белки могут создавать препятствия для компонентов
белоксинтезирующего аппарата (мешать связыванию с мРНК или ее сканированию).
При связывании с мРНК на большом расстоянии от места инициации регуляторные
белки могут влиять на процесс инициации путем изменения общей
пространственной структуры мРНК, изменяя таким образом доступность
инициирующего кодона или 5'-конца мРНК для белоксинтезирующего аппарата. Регуляторными белками могут быть специальные белки клетки,
выполняющие только эту функцию в организме, а также белки, имеющие в
организме другие функции и работающие в качестве регуляторных белков "по
совместительству". Довольно часто в качестве белков, регулирующих
активность определенных мРНК, могут выступать сами продукты их трансляции
(авторегуляция)».
[1]. Нормальная ситуация. Если не хочешь, чтобы это читали –
поставь белковое заграждение и знак – «Объезд». И считывающая машинка пройдет
мимо. Потому, сначала сканируем всю мРНК, читаем правила,
узнаем, какие дополнительные команды нужны, вносим коррективы… и только тогда
трансляция идет, как надо. Даже для мРНК вполне стандартного вида. Процессиг мРНК идет долго. Для него необходимо место и
время. Кстати, именно по этой причине в эукариотических клетках процессы
транскрипции и трансляции не только разделены по времени, но и разнесены в
разные области клетки. И, мало того, перед отправкой сформированной
стандартной мРНК из ядра клетки в цитоплазму она проходит проверочный тест.
Похоже, что новая мРНК проходит пробную трансляцию. И только в том случае, если она исправно работает,
белок получен, только тогда мРНК отправляется в рабочую область
трансляции. Далее еще некоторые видоизменения чтения, видимо, уже
для прокариотов: «Некоторые мРНК
содержат сигналы на изменение рамки считывания. Некоторые мРНК содержат в
транслируемой области терминирующие кодоны, но эти кодоны успешно обходятся
за счет изменения рамки считывания перед ними или непосредственно на них.
Рамка может сдвигаться на -1, +1 и + 2. Существуют специальные сигналы в
мРНК, изменяющие рамку считывания. Так, сдвиг рамки трансляции на -1 на РНК
ретровируса происходит на специфической гептануклеотидной последовательности
перед шпилечной структурой в мРНК (рис. 5, в). Для сдвига рамки на +1 на мРНК бактериального
фактора терминацинации RF-2 важны нуклеотидная
последовательность на месте сдвига (кодон UGA), последующий кодон, а также предшествующая им последовательность,
комплементарная к 3'-концевой последовательности рибосомной РНК (аналог последовательности
Шайна-Дальгарно) (рис. 5, г)». [1]. Даже наши современные компьютеры не могут так лихо
перестраивать головки считывания информации с носителей. Да, надо признать,
что техника считывания информации с мРНК очень серьезная. Тут эволюция постаралась.
Продолжим смотреть особенности считывания. Дальше не
менее интересно: «Считывание мРНК в пределах
одного цистрона не всегда является непрерывным. Первоначально считалось, что
последовательность нуклеотидов в мРНК всегда читается непрерывно от
инициирующего до терминирующего кодона. Однако оказалось, что при трансляции
мРНК гена 60 фага Т4 последовательность значительной длины может пропускаться
(рис. 5, д). При этом рибосома совершает
как бы прыжок по мРНК с одного глицинового кодона GGA, находящегося перед терминирующим кодоном UAG, на другой глициновый кодон GGA,
который отстоит от первого на 50 нуклеотидов. Механизм этого явления пока не
очень ясен. Не исключено, что такое шунтирование мРНК обеспечивается ее
особой пространственной структурой, например выпетливанием той части молекулы
мРНК, которая пропускается при трансляции». [1] Подводим итог: «Все приведенные примеры нарушения общих правил
кодирования так или иначе связаны с существованием определенного контекста в
мРНК. Этот контекст или перекодирующие сигналы иногда называют вторым
генетическим кодом». [1] Очень важный вывод. Алгоритм выполнения трансляции,
заложенный в интронах, не менее важен, чем механизм экзонов - исполнительной
части задачи трансляции. И очень возможно, что второй генетический код может
скоро стать первым, по степени важности в клеточных процессах. По мере его
понимания… Нам осталось только констатировать: в формировании и чтении эукариотической мРНК развитие
техники считывания информации шло по пути стандартизации формы (введение
КЭП, 5', 3', поли А -
последовательности), а в технике чтения прокариотической мРНК больше
развивался читающий автомат - рибосома. И потому эти процессы надо изучать
раздельно. Иначе только лишняя путаница возникает. Рибосома.
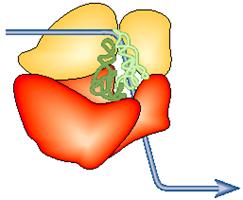
Вот он, уникальный считывающий автомат, который
превращает последовательность нуклеотидов мРНК в белок, или копию, или набор
химический команд… Рибосома должна учитывать все хитрости считывания
информации: менять последовательность чтения, сдвигать рамку считывания,
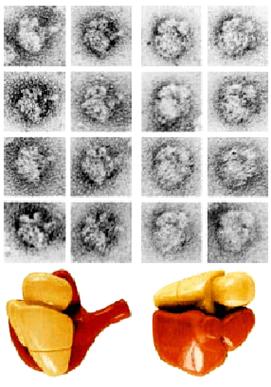
менять смысл прочитанного триплета в соответствии с контекстом… « Рибосома играет роль организующего
центра в чтении генетической информации. Это молекулярная машина, построенная
по единой схеме у всех организмов с некоторыми вариациями. Она состоит из
двух рибонуклеопротеидных субчастиц: малой и большой. В состав малой
субчастицы у бактерий, например, входит молекула рибосомной РНК (рРНК) длиной
~1500 нуклеотидов плюс еще одна небольшая молекула рРНК (110-120 нуклеотидов)
и 21 разный белок. В состав большой субчастицы - одна молекула рРНК длиной ~
2700 нуклеотидов плюс 32 разных белка. Если каждый белок рибосом обычно
закодирован в одном гене, то для кодирования рРНК характерна большая генная
избыточность. Так, например, у кишечной палочки каждую молекулу рРНК кодируют
7 генов, а у дрожжей - несколько сот генов. «Физически рибосома представляет собой компактную
частицу специфической формы, лишенную внутренней и внешней симметрии, грубо
аппроксимируемую сферой с диаметром около 30 нм. Функционально это
молекулярная машина, протягивающая вдоль себя цепь мРНК, считывающая
закодированную в мРНК генетическую информацию и параллельно, в соответствии с
кодом, синтезирующая полипептидную цепь белка из поступающих в нее
аминокислотных остатков. В процессе работы рибосома потребляет энергию гидролиза гуанозинтрифосфата (ГТФ). Очевидно, что детальное
знание структуры рибосомы является необходимой базой для понимания механизмов
работы этой молекулярной машины.
Подумаем…
Теперь попробуем осмыслить процесс считывания
информации с ДНК и вставить недостающие звенья. Наиболее четко процесс
считывания информации прослеживается в эукариотическтх клетках. Но, сначала некоторые общие моменты… Клетка не может с нуля создать самое себя. Сложность
воспроизводства всей системы настолько сложна, что повторить её силами клетки
невозможно. Её можно только копировать, воспроизводя то, что уже накоплено.
По частям, постепенно создавая копию. Даже не копию, а часть копии. Полностью
копируется только информация хромосом клетки. И центр копирования – ядро.
Остальное только частично копируется и доводится до работоспособности. И
только время жизни ядра определяет жизненный цикл клетки. Возраст клетки
определяет состояние её логического центра управления. Всё остальное
ремонтируется. Вот тут заметим, что новая клетка образуется из части
старой клетки, деля внутреннее пространство материнской клетки на свою сферу
управления и старую, воспроизводящую.
Сначала функционально, а потом и фактически. И понятно, почему. Команды управления – химические.
Канал передачи информации – внутриклеточная жидкость. Я бы отметил максимальность в режимах всех клеточных
процессов. Она заложена изначально. Потому, что это - целевая задача клетки.
Она не может ограничиваться. Но, постоянное наращивание мощности всех
производственных потоков постепенно приводит к перегрузкам в работе центра
управления - ядра. В какой-то момент оно уже не справляется с нагрузкой…,
надо усиливать каналы управления, а программа их усиления только одна –
копирование. И начинает создаваться дублирующий центр управления. Пока, как
вспомогательный. По мере строительства новый центр осваивает и дублирует
все каналы управления. Когда вновь создаваемый управляющий центр клетки
начинает работать самостоятельно и вмешиваться в работу оригинала, то
возникает перекрытие зон управления. Но, постепенно каждое ядро формирует
свою зону управляемости, комплектует свой необходимый набор составляющих
полного автономного цикла, не обращая внимания на то, свой это объект или
нет. И возникает конкуренция центров
управления. В этой ситуации надо делить сферы управляемости не только по
функциональному набору, но и фактически…, потому, что канал передачи
информации общий – внутриклеточная жидкость. Возникает необходимость ставить перегородки, чтобы
отделить свою область управления. Это делают оба центра управления. И, таким
образом заканчивают деление клетки на два пространства управления. Вот теперь начинается осмотр доставшегося при дележе
имущества. Комплектность проверялась чуть раньше, на стадии раздела сфер
управляемости, теперь проводится только ревизия доставшегося. Чего и сколько
не хватает для полноценной деятельности. Центры управления обеих половинок
снова работают в полную силу, доводя производственные линии всех процессов
клетки до максимальной мощности. Этот процесс бесконечен… Но, вернемся к процессу считывания информации. И снова
вопросы… Как идет обмен информацией внутри клетки? Химическими соединениями. Других вариантов
вроде нет. Что это за соединения такие? Ферменты, гормоны, блокаторы,
активаторы, разные РНК,… примерно это. Где производятся химические команды? Их производят
рибосомы. В ядрышке на основе
имеющейся там РНК, в ядре на той же основе ядерной РНК. Эти команды поступают
во внутриклеточную жидкость и доходят до адресата. Там принимаются, пусть
пока теми же рибосомами. И рибосомы, например, уже в цитоплазме начинают их
исполнять. Производить белок, считывать информацию с мРНК … и т.д. Процесс
управления тут как-то понятен. Обратная связь работает по тому же принципу. Ответы
центру управления формируют снова рибосомы. Примерно такими же химическими
соединениями, но их принять и понять может только центр управления – ядрышко.
Конечно, видов обратной связи в клеточных процессах
много и разных, но нас пока интересует только этот. От ядрышка к ядерным
рибосомам, от ядерных рибосом к рибосомам в цитоплазме и обратно к ядерным рибосомам и ядрышку.
Эти каналы химической связи контролируют копирование ДНК, создание
РНК-предшественника и стандарной мРНК, производство белков, а так же и цикличность деления клетки. И на всех значимых контрольных пунктах работают
рибосомы – логические машины клетки.
Их много и они разные. Каждая четко оринтирована на выполнение и
контроль только своей задачи. Она работает на основе поступающих команд,
определяет скорость и условия выполнения работы, создает отчеты о выполнении
и ответные команды управления к центру. Уникальная машина. Единственный
автомат в клетке, который в состоянии воспроизвести самое себя. С нуля. Рибосомы в клетке производят всё: РНК, мРНК, тРНК,
рРНК, белки, самих себя, команды управления…. Все первичные функции воспроизводства
отданы рибосомам. От информации до строительных материалов. Эволюция клетки
определяется эволюцией рибосом. И с этой стороны ядрышко ядра клетки вполне
может оказаться или конгломератом рибосом или одной большой рибосомой с
набором уникальных функций. А в основе работы этого уникального автомата лежит
считывание информации с РНК. РНК – это, сначала, всё же, программа,
записанная на стандартном носителе в стандартных единицах системы. А потом уже транспортное средство и
остальные её применения. Внутри каждой рибосомы такая программа есть - рРНК. Вот она и определяет все процессы. Внутри рибосомы проиходит то же самое, что
и снаружи. Чтение РНК. С одной разницей: внутреннее чтение имеет целью
управления внутренними процессами и контролем за выполнением внешних. Скорее всего, многообразие выполняемых рибосомами
функций и обеспечивается соответствующим набором необходимых рРНК. Какая рРНК вложена в рибосому, такая
программа и начинает выполняться этой, похоже, вполне стандартизованной
машиной. Правда, как это происходит, видимо, пока не знает
никто… Пример устройства и организации рибосомы…, с этим
сейчас уже трудно, это из прошлого нашей вычислительной техники. Когда
исходная и конечная информация еще находились на перфолентах, длинных узких бумажных лентах с пробитыми отверстиями
в нужных местах. Представим себе такую программируемую машину. Одна
такая перфолента (рРНК), внутри
машины. На ленте записана программа действий. Начало и конец рабочих тактов
считывания части информации с неё задается внешними сигналами на соответствующие
входы. Пришел стартовый сигнал – лента начала двигаться и с неё считывается
программа. И исполняется. Пришел сигнал СТОП – лента остановилась. Машина ждет дальнейших команд. Снова пришел
стартовый сигнал – работа возобновилась … и снова СТОП… Но, разных управляющих входов в такой считывающей
машине много, и работа, даже по простой последовательной программе, может
очень сильно меняться в зависимости от этого внешнего управления. Хорошо, машина работает по программе, а что же она
делает? Она выполняет копирование еще одной перфоленты, внешней, вложенной во
внешнее считывающее устройство. Исходный оригинал – перфолента (мРНК), а вот
какую копию мы получим на выходе, зависит от внутренней программы (рРНК). В
одном случае такую же перфоленту (мРНК или рРНК), в другом - сборную строительную конструкцию (белок),
в третьем – транспортное средство
(тРНК)…. В зависимости от состояния
разных входных сигналов работа синхронизируется с другими устройствами,
возникает разнообразие конструкций и размеров, и т.д. Но, в общем случае,
машина выполняет одну и ту же работу –
копирует исходный оригинал, только вот результат во всех случаях – разный.
Такая вот универсальная машина – рибосома. Теперь коснемся самой информации. Информация ДНК и РНК – противоположности. Абсолютные.
Потому, что получаются они контактным копированием. Как позитив и негатив. И
чтобы получить полную копию информации ДНК, необходимо процесс копирования
повторить дважды. Позитив – негатив - позитив. Таблицы соответствия аминокислот тем или иным триплетам
ДНК и РНК [1,2] убеждают нас, что триплеты почти идентичны (Т меняется U).
Каким образом они получились такими, видимо, специалистам понятно, а вот нам
дилетантам – не очень. Так как, при контактном копировании между ДНК и мРНК должна
существовать еще одна, промежуточная копия РНК - негативная, по отношению к
этим. Но, как мы видели, в описании процессов считывания [1,2], об этом нет
ни слова. И триплеты соответствия на тРНК должны быть негативны
по отношению триплетов считывания на мРНК, т.е. иметь противоположный код.
Этот момент учтен [2,3], но не введен, как правило копирования. Если эти мелочи выкинуты из учебников для создания
простоты понимания, то хотя бы одной строкой в научных работах они должны
присутствовать и учитываться при описании процессов считывания информации ДНК
и формирования её копии. В работе [9] уже указывалось на невозможность
использования только неперекрываемого триплетного кода при считывании
информации с ДНК и мРНК. Нет четкой фиксации триплетов. Неперекрываемый код
возможен только для высокоразвитых рибосом с двойным контролем считывания,
как частный случай, и никак не может быть всеобщим. Но генетикам видимо не до этого… Заключение.
Не думаю, что в этой статье сказано что-то такое, что
неведомо специалистам. Изучение внутриклеточных процессов на уровне
математики [4], и уж тем более, таких, как информация ДНК, идет очень
активно. По математической оценке биохимических процессов получения
информации ДНК материалов не меньше, чем по клетке в целом. О генетических
исследованиях написано еще больше. Тем более странно, что я не вижу материалов о
рассмотрения процессов в клетке с
этой, технической стороны. Вполне очевидной, если вдуматься. Возможно, что я просто не знаю, что и где
смотреть. Если читатели знают о работах и исследованиях в этом направлении,
опубликованных ранее, то прошу сообщить.
Екатеринбург
andvnikitin@yandex.ru Октябрь 2010г. Литература:
1.
Л. П. ОВЧИННИКОВ Что и как закодировано в мРНК. Московский
государственный университет им. М.В. Ломоносова http://nature.web.ru/db/msg.html?mid=1157645&s= 2.
С. Г. ИНГЕ-ВЕЧТОМОВ Трансляция как способ существования живых систем, или
в чем смысл "бессмысленных" кодонов. Санкт-Петербургский
государственный университет. http://nature.web.ru/db/msg.html?mid=1157633&s= 3.
А. С. СПИРИН Принципы структуры рибосом. Московский государственный
университет им. М.В. Ломоносова http://nature.web.ru/db/msg.html?mid=1157656&s 4.
Г. Р. ИВАНИЦКИЙ, В. И. КРИНСКИЙ, Е. Е. СЕЛЬКОВ. МАТЕМАТИЧЕСКАЯ БИОФИЗИКА
КЛЕТКИ. ИЗДАТЕЛЬСТВО «НАУКА» * МОСКВА 1978.
http://www.y10k.ru/books/detail7152.html
5.
База знаний по геронтологии. http://gerontology-explorer.narod.ru/2c90c098-9b2b-4f70-b34d-67a5f4c0d428.html 6.
Эукариоты.. Википедия, свободная энциклопедия. http://ru.wikipedia.org/?oldid=28231750 7.
Интрон. Википедия, свободная энциклопедия. http://ru.wikipedia.org/?oldid=27905411 8.
Сплайсинг. Википедия, свободная энциклопедия. http://ru.wikipedia.org/?oldid=28272571. 9.
А. В. Никитин Триплеты в ДНК. http://trinitas.ru/rus/doc/0016/001c/00161697.htm 10.
А.В.Никитин Эволюционный
путь саморазвития искусственного интеллекта.
http://andrejnikitin.narod.ru/samorazvitiemozg2.htm |
||||||||||
|
|